
Ощущения рецепторы которых расположены в связках
Содержание статьи
Биология в лицее
Осязание. Мышечное чувство и двигательный анализатор Осязание — это способность организма воспринимать различные воздействия внешней среды и преобразовывать поступающие сигналы в определённый вид чувствительности. В осязании выделяют ощущения прикосновения и давления (тактильная чувствительность), тепла и холода (температурная чувствительность, или терморецепция), боли (ноцицептивная чувствительность) и другие смешанные ощущения. Органами осязания являются рецепторы, находящиеся в наружном покрове, мышцах, сухожилиях, суставах, некоторых слизистых оболочках (губ, языка, половых органов).
Значение осязания для организма очень велико. Чувство боли предохраняет организм от травм, ожогов, обморожения, сообщает о возникновении заболевания. Ощущение давления помогает нам ориентироваться во время ходьбы и бега. Одни рецепторы кожи воспринимают холод, другие — тепло, третьи — давление, четвертые — прикосновение и т. д. Для того чтобы убедиться, что рецепторы прикосновения и холода различны, достаточно провести по тыльной стороне руки остро заточенным карандашом. Грифель холоднее кожи (температура комнаты обычно около 20 °С, а температура кожи ладони около 36 °С). Возбуждение тактильных рецепторов даст нам возможность почувствовать скольжение карандаша, а рецепторы, воспринимающие холод, будут лишь периодически сообщать об изменении температуры кожи в местах прикосновения, да и то лишь в том случае, если мы наткнемся на точку холода, т.е. на то место, где располагаются эти рецепторы. Рецепторы тепла обнаружить несколько сложнее. Во-первых, потому что острие придется нагреть примерно до 40 °С (выше не надо, так как горячий предмет вызывает боль). Во-вторых, потому что точек, воспринимающих тепло, меньше. Так, на 1 см2 кожи кисти приходится 1-5 холодовых точек и только 0,4 тепловых.
Это чувство возникает потому, что в мышцах, сухожилиях, связках и суставах находятся рецепторы, которые называются проприорецепторами. Они возбуждаются при мышечных сокращениях. Проприорецепторы посылают информацию в мозг о положении частей тела в пространстве, о состоянии мышц, о степени их напряжения. Благодаря их деятельности ЦНС получает непрерывные сигналы о состоянии скелетной мускулатуры, в результате чего может осуществляться непрерывная сложнокоординированная двигательная деятельность организма. Ядро проприоцептивной чувствительности залегает в задней центральной (постцентральной) извилине теменной доли коры.
Двигательный анализатор — это анализатор, который даёт представление о положении тела в пространстве и его отдельных частей, о степени сокращения мышц, натяжении связок. С помощью этого анализатора человек знает о положении своего тела и без зрительного контроля — при закрытых глазах.
Возбуждение, возникшее в рецепторах двигательного анализатора, по центростремительным нервам через задние (чувствительные) корешки проводится в спинной мозг. По восходящим проводящим путям оно передаётся в кору головного мозга. Проводниковый отдел этого анализатора передаёт импульсы также в другие центры головного мозга: продолговатый и средний мозг, мозжечок. Центральная часть двигательного анализатора — это двигательная зона коры головного мозга, а именно передняя центральная (предцентральная) извилина, расположенная в лобной доле полушарий большого мозга.
Для отражения относительных размеров областей представительства отдельных участков тела в коре предцентральной извилины полушарий большого мозга Пенфилдом и Расмуссеном создано изображение двигательного «гомункулюса». Как видно из этого изображения, наибольшее представительство в коре имеют кисть и пальцы, а также мышцы головы, в основном мимические. Двигательный анализатор взаимодействует с другими анализаторами, например со зрительным и слуховым анализаторами. Развитие трудовой деятельности привело к тому, что наибольшей тактильной чувствительностью обладают пальцы рук. Большая осязательная чувствительность губ связана с поступлением пищи в организм. < Предыдущая страница «Органы слуха и равновесия. Их анализаторы» Следующая страница «Органы обоняния и вкуса» > | Календарь
СтатистикаОнлайн всего: 1 Гостей: 1 Пользователей: 0 | |||||||||||||||||||||||||||||||||||||||||||||
 Осязание дает представление о поверхности предмета, его форме, размерах, массе. Когда мы касаемся предмета, держим его или ощупываем, в нервных окончаниях кожи, а также рецепторах мышц и сухожилий возникает возбуждение. Возбуждение по нервам передаётся в головной мозг — в зону кожно-мышечной чувствительности теменной доли коры больших полушарий головного мозга. Возникают ощущения массы предмета, состояния его поверхности.
Осязание дает представление о поверхности предмета, его форме, размерах, массе. Когда мы касаемся предмета, держим его или ощупываем, в нервных окончаниях кожи, а также рецепторах мышц и сухожилий возникает возбуждение. Возбуждение по нервам передаётся в головной мозг — в зону кожно-мышечной чувствительности теменной доли коры больших полушарий головного мозга. Возникают ощущения массы предмета, состояния его поверхности.
 Мышечное чувство — это процесс восприятия раздражений, возникающих при изменении степени сокращения и расслабления мышц.
Мышечное чувство — это процесс восприятия раздражений, возникающих при изменении степени сокращения и расслабления мышц.
 Периферической частью двигательного анализатора служат внутренние рецепторы органов движения — мышц, суставов и сухожилий. Они получают раздражения во время движения этих органов и, посылая импульсы в кору полушарий, сообщают о состоянии органов движения и о тех действиях, которые человек совершает с их помощью.
Периферической частью двигательного анализатора служат внутренние рецепторы органов движения — мышц, суставов и сухожилий. Они получают раздражения во время движения этих органов и, посылая импульсы в кору полушарий, сообщают о состоянии органов движения и о тех действиях, которые человек совершает с их помощью. В это место поступают импульсы со всех других чувствительных зон коры полушарий большого мозга, поэтому передняя центральная извилина является «центральным аппаратом построения движений» и обеспечивает формирование целенаправленных реакций в ответ на внешние стимулы.
В это место поступают импульсы со всех других чувствительных зон коры полушарий большого мозга, поэтому передняя центральная извилина является «центральным аппаратом построения движений» и обеспечивает формирование целенаправленных реакций в ответ на внешние стимулы.Источник
Рецепторы скелетных мышц и суставов человека
Во второй лекции по дисциплине «Биомеханика мышц» для студентов НГУ им. П.Ф.Лесгафта описывается состав и строение мышечных веретен — рецепторов, расположенных в мышцах и информирующих нервную систему о текущей длине мышцы и скорости ее сокращения, и сухожильных рецепторах Гольджи, которые информируют нервную систему о степени возбуждения мышцы. Также в лекции описываются уровни построения движений по Бернштейну и их взаимосвязь с функционированием рецепторами опорно-двигательного аппарата.
Лекция 2
Функционирование рецепторного аппарата мышц
2.1. Рецепторы опорно-двигательного аппарата человека
При изучении анатомии и физиологии (А.С. Солодков, Е.Б. Сологуб, 2001) вы изучали двигательную сенсорную систему. Одним из отделов этой системы являются проприорецепторы, расположенные в мышцах, сухожилиях и суставных сумках. В мышцах расположены мышечные веретена, в сухожилиях — сухожильные органы Гольджи. В суставных сумках расположены рецепторы суставов.
Мышечные веретена
Еще в XIX веке В. Кюне обнаружил в скелетных мышцах структуры, напоминающие веретено. Затем, в начале XX века Нобелевский лауреат Чарльз Скотт Шеррингтон показал, что эти структуры служат чувствительными рецепторами. Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Интрафузальные волокна подразделяются на два типа:
- Длинные и толстые (диаметр 20-25 мкм)[1], которые информируют ЦНС о динамическом компоненте движения — скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
- Короткие и тонкие (диаметр 10-12 мкм), которые информируют ЦНС о статическом компоненте движения — текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
Рекомендую обратить внимание на учебные пособия «Биомеханика мышц» и «Гипертрофия скелетных мышц человека«
Рецепторы Гольджи
Нервно-сухожильные веретена (рецепторы Гольджи) открыл в 1903 году Камилло Гольджи. Впоследствии за эти исследования ему была присуждена Нобелевская премия. Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр — 0,1-0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна сухожилия натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис.2.1). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 — 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
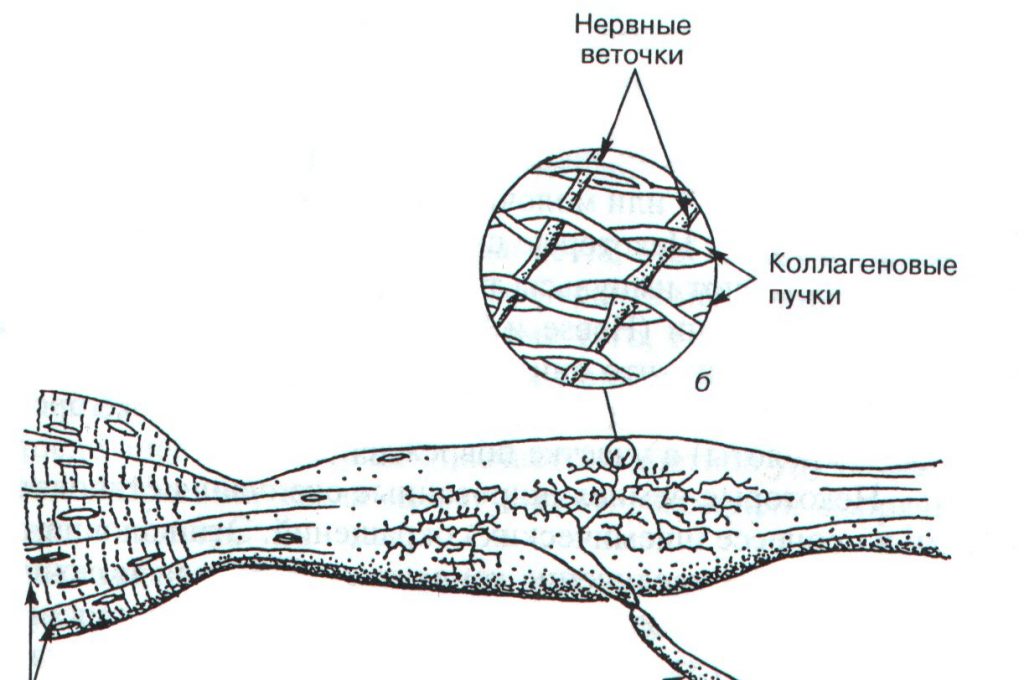
Рис. 2.1. Строение сухожильного органа Гольджи (А.Дж. Мак-Комас, 2001)
Рецепторы суставов
Суставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
2.2. Уровни построения движений и рецепторы опорно-двигательного аппарата
В 1947 году была опубликована книга русского ученого Н.А. Бернштейна «О построении движений». В этой книге он изложил оригинальную теорию управления двигательными действиями. В основе этой теории лежало понятие уровней построения движений. Он писал: «Каждая двигательная задача находит себе в зависимости от своего содержания и смысловой структуры тот или иной уровень. Иначе говоря, тот или иной сензорный синтез, который наиболее адекватен по качеству и составу образующих его афферентаций и по принципу их синтетического объединения требующемуся решению этой задачи. Этот уровень и определяется как ведущий уровень для данного движения» (Н.А.Бернштейн, 1991, С.41). По концепции Н.А. Бернштейна ни одно движение не обслуживается только одним уровнем, но осознается только ведущий уровень. По классификации Н.А.Бернштейна, при управлении движениями может быть задействовано пять уровней. Проприорецепторы опорно-двигательного аппарата человека составляют основу двух нижних уровней: уровня А и В.
Афферентация уровня А основана на импульсации мышечных веретен (длина и скорость сокращения мышцы) и рецепторов Гольджи (уровень возбуждения мышцы при ее укорочении). Эта информация очень слабо осознается ЦНС, то есть, по гипотезе Н.А.Бернштейна, этот уровень почти никогда не бывает ведущим.
Афферентация уровня В опирается на информацию, поступающую от суставных рецепторов. Это уровень выступает как ведущий в ряде физических упражнений, таких как наклоны тела вперед и назад, а также циклические движения. Сигналы от суставных рецепторов хорошо осознаются.
Рекомендуемая литература
1. Самсонова, А.В. Гипертрофия скелетных мышц человека: учебное пособие /А.В.Самсонова, СПб: Кинетика.- 159 с., ил.
2. Самсонова А.В. Биомеханика мышц: учебно-методическое пособие /А.В.Самсонова, Е.Н. Комиссарова / Под ред А.В.Самсоновой .- СПб., 2008.- 217 с.
3. Самсонова А.В. Моторная и сенсорная функции мышц в биомеханике локомоций: монография /А.В.Самсонова; СПбГУФК им. П.Ф.Лесгафта, 2007.- 152 с.
Источник
XIII. ()
, , . . , . . (), (), , , , , . , . . , , , . . : . , (, , ). , , . . . , , . . . , , . . — — , , . , . . . . . , . — , , . . , . . . . . , , . . — , . , , . , , . , . . , . . () — , 100 . . 59 , — 70,5 . , . , . . , . , . , . , , (. 91). , , . . , , , .
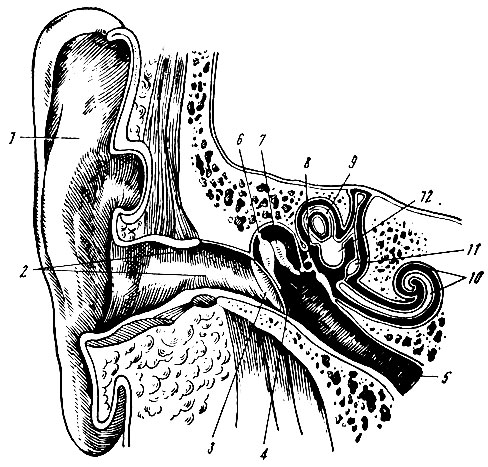
. 91. ( ). — , — . — . , , , (. 92).
. 92. (1), (2) (3) . , , , . , . . , . , . . , , , . , — . . , , . . . — — . , , , , , . , , — . . , . . . , , , . . . . , 4 . 6-7 . 110-125 . . . ( 140 . 1 2). , . . , , . — () — . . . , , , . 93, ( 25 ) , .
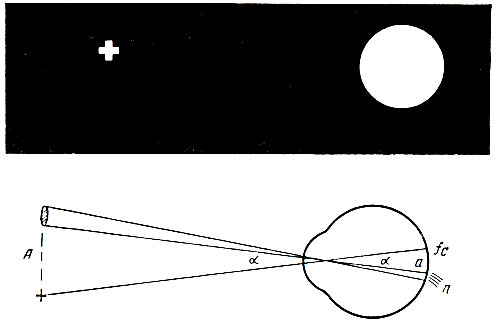
. 93. . . , , (n), — (fc) . , . , , . . , . — . , , , . 17, 18 19- . , — , , . . . . , . . 15 . . , , , . . . . . — , . , , , , . , «» , . , . . . . , , — . . . (1756) , . . . , . , , , — . , , . , , , . . . 620-760 (6200-7600 ) , 510-550 (5100-5500 ) — , 390-450 (3900-4500 ) — . , , , , , . . . , . , . . , , . . . , , , . . . . . , , . . , . . — , , (. 94). — — , . , .
. 94. . 1 — ; 2 — ; 3 — ; 4 — ; 5 — ; 6 — ; 7 — ; 8 — ; 9 — ; 10 — ; 11 — ; 12 — , , , , . , . . , , , , . — () , . , . , , . . () , , , . . , . — , . : , , , , (. 95). . . : , (. . 96).
. 95. ( ). 1 — ; 2 — ; 3 — ; 4 — ; 5 — ; 6 — ; 7 — ; 8 — ; 9 — ; 10 — ; 11 — ; 12 — ; 13 — ,
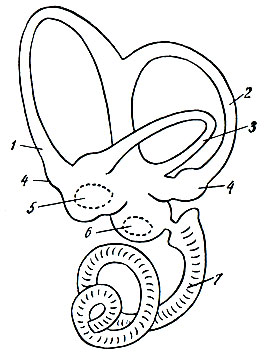
. 96. . 1, 2, 3 — ; 4 — ; 5, 6 — ; 5 — ; 6 — ( ); 7 — , . . . , . , , . . . . . — . , , . , — . , . . . . . — . . , . : , . . . . , , . . , . , . — 16 20000 1 . . , , . , , . , . — . , . , . , , . . . — , . , , , — , — . (. 96). — () (). , — . , . . . . . , , . . . , . . , . . . . . , , , , , . — . (), (), ( ). () : — , , — , . , , , , , . , , . . . , , , — ( ), — . —, , , , . : , ; , ; , , , . , . , ( ) . . , , -. , , , . . -. , , . , — — , — . , . . . , . . , , , . . ( 2 6) . . — . . . , , , , . . , , . , . , , , — . . , . . , , — . , . . ( ), ( ) , , . , , , . «», «», «» . . . , . . , . . , , 40 , . . , . , , . . . . . , , . ,. . , , , . , , , , , . , , . . , , , , , , . , . , . , , . : , , , . , , , . , . , . , — . . : , — . , . , , . , . . , . , , , , , . , , . , . , . , , , XII ( ). . , , . . . , (), (), () (). , . . , , . |

Источник
Физа / Физа / Занятия / Разное / ПРОПРИОЦЕПТИВНЫЙ АНАЛИЗАТОР
проприоцептивныйАНАЛИЗАТОР
Систематическая классификация была предложена английским физиологом Ч. Шеррингтоном (1857-1952). Взяв за основу характер отражения и место расположения рецепторов, он разделил все ощущения на три группы: экстероцептивные, проприоцептивные и интероцептивные.
Общая характеристика.
Двигательный анализатор играет важную роль в адаптации организма к изменяющимся условиям окружающей среды. Наряду с кожным, зрительным, вестибулярным двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности
В 1826 году Бенон высказал предположение, что мышечная система — не только рабочий орган, но и своеобразный орган чувств, работа которого приводит к возникновению мышечного чувства.
По мнению И.М. Сеченова (1863) мышечное чувство, т.е. чувство положения тела в пространстве, возникает при активации специфических рецепторов мышечной системы. . «тёмным мышечным чувством». Эти рецепторы Ч. Шеррингтон назвал проприорецепторами, т.е. собственными рецепторами опорно-двигательного аппарата (лат. proprius — собственный).
Проприорецепция (глубокая, или кинестетическая чувствительность) является разновидностью интерорецепции. Она представляет собой восприятие позы и движения нашего собственного тела. Поза определяется улом расположения костей в каждом суставе, устанавливаемым либо пассивно (внешними силами), либо активно (мышечным сокращением). С их работой сочетаются сигналы от вестибулярного органа, что позволяет определять положение тела в поле земного притяжения. Проприорецепторы участвуют также в нашей сознательной и бессознательной двигательной активности.
Таким образом, проптиорецепторы — это группа тканевых, первичночувствующих, механорецепторов, обуславливающих поступление информации о положении тела в пространстве и о взаиморасположении частей, отделов тала относительно друг друга. Все эти рецепторы представляют собой механорецепторы, специфическим раздражителем которых является их растяжение
Проприорецепторы располагаются в суставах, мышцах, сухожилиях и связках, надкостнице, коже. Они возбуждаются при деятельности опорно-двигательного аппарата, участвуя в анализе сигналов, поступающих от его элементов, а затем передают информацию в ЦНС. От 30 до 50% нервов, идущих к мышцам, составляют нервные волокна, несущие импульсы от проприорецепторов мышечного аппарата.
Рефлексы с проприорецепторов замыкаются на различных уровнях ЦНС и обуславливают мышечный тонус, позу и координацию движений. Благодаря наличию скелетных мышц мы можем взаимодействовать с окружающей средой. Эти мышцы позволяют нам осуществлять самые разнообразные движения: бег, ходьбу, речь, письмо, мимику, жесты.
Для правильного осуществления всех этих движений необходимо, чтобы в центры движения постоянно поступала информация о положении тела в пространстве, и о том, как совершается движение. Поэтому двигательный анализатор не способен к адаптации.
Проприорецепция — способность мгновенной оценки и контроля положения тела в пространстве (без участия зрения) — имеет исключительное значение в спорте, а в футболе, с его сложнейшей техникой, в особенности.
Именно в футболе нередки ситуации, когда спортсмен выполняет сложные по координации движения без зрительного контроля.
При расстройствах проприоцепции возникают сенситивная атаксия и псевдоатетоз. Так, описан (Витгенштейн) пример потери проприоцепции после операции, что привело к тому, что пациентка постепенно разучилась двигаться, есть и даже переставала дышать. Возвратить к жизни её удалось путём замены проприоцепции другими чувствами, в первую очередь — для движения — зрением.
Двигательный (проприоцептивный) анализатор обеспечивает формирование так называемого «мышечного чувства» при изменении напряжения мышц, их оболочек, суставных сумок, связок, сухожилий. В мышечном чувстве можно выделить три составляющих:
чувство положения, когда человек может определить положение своих конечностей и их частей относительно друг друга;
чувство движения,- это информация о направлении и скорости движения суставов. Человек воспринимает как активное движение сустава при мышечном сокращении, так и пассивное, вызванное внешними причинами. Порог восприятия движения зависит от амплитуды и от скорости изменения угла сгибания суставов.
чувство силы, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определенном положении при подъеме или перемещении груза..
Периферический отдел представлен проприорецепторами, расположенными в мышцах, связках, сухожилиях, суставных сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца Фатера-Пачини, свободные нервные окончания.
Мышечное веретенопредставляет собой скопление тонких коротких поперечнополосатых мышечных волокон, которые окружены соединительнотканной капсулой. Эти волокна получили название интрафузальных в отличие от обычных мышечных волокон, которые составляют основную массу мышц и называются экстрафузальными, или рабочими, волокнами. Интрафузальные волокна расположены параллельно экстрафузальным, поэтому возбуждаются при расслаблении (удлинении) скелетной мышцы.Строение мышечного веретена. Мышечное веретено длиной 3 мм шириной 0,3 мм состоит из интрафузальных мышечных волокон, которые окружены толстой соединительнотканной капсулой, внутри которого находится жидкость, подобная лимфе. Два толстых волокна имеют по экватору скопление ядер (ядерная сумка), и волокна называются ядерно-сумчатыми. Четыре тонких волокна имеют по экватору ядра расположенные в один ряд (ядерная цепочка), а волокна получили название ядерно-цепочечных.
В разных скелетных мышцах число веретен на 1 г ткани варьирует от нескольких единиц до сотни.
. Мышечные веретена прикрепляются к мышечным волокнам параллельно-один конец к сухожилию, а другой-к волокну. Каждое веретено покрыто капсулой, образованной несколькими слоями клеток, которая в центральной части расширяется и образует ядерную сумку. Внутри веретена содержится несколько (от 2 до 14) тонких внутриверетенных или так называемых интрафузальных мышечных волокон. Эти волокна в 2-3 раза тоньше обычных волокон скелетных мышц (экстрафузальных).
Интрафузалъные волокна подразделяются на два типа: 1) длинные, толстые, с ядрами в ядерной сумке, которые связанны с наиболее толстыми и быстропроводящими афферентными нервными волокнами — они информируют о динамическом компоненте движения (скорости изменения длинымышцы) и 2) короткие, тонкие, с ядрами, вытянутыми в цепочку, информирующие о статическом компоненте (удерживаемой в данный момент длине мышцы).
Окончания афферентных нервных волокон намотаны на интрафузальные волокна рецептора. При растяжении скелетной мышцы происходит растяжение и мышечных рецепторов, которое деформирует окончания нервных волокон и вызывает появление в них нервных импульсов. Частота проприоцептивной импульсации возрастает с увеличением растяжения мышцы, а также при увеличении скорости ее растяжения. Тем самым нервные центры информируются о скорости растяжения мышцы и ее длине. Вследствие малой адаптации импульсация от мышечных веретен продолжается в течение всего периода поддержания растянутого состояния, что обеспечивает постоянную осведомленность центров о длине мышцы
Рассмотрим подробно миотатический рефлекс. В мышце есть три основных типа проприорецепторов:
1) первичные окончания веретён;
2) вторичные окончания веретён;
3) рецепторы Гольджи.
На интрафузальных волокнах спирально расположены чувствительные окончания афферентных волокон группы Iа — Эти волокна называются первичными афферентами. Первичные окончания(около 15 мкм) -миелинизированное чувствительное нервное волокно Ia (скорость проведения 70-120 м/с) формирует первичное, или аннулоспиральное окончание, (они представляют из себя рецепторную спираль). Эти окончания возбуждаются при расслаблении или удлинении мышцы, вызванной растяжением, и реагируют на скорость удлинения экстрафузальных волокон.
При механической деформации аннулоспиральных окончаний (периферическое воздействие) рождается рецепторный потенциал, который далее преобразуется в потенциал действия и распространяется по толстым афферентным волокнам первой группы (Iа) к спинальным ганглиям. Далее волокна от спинального ганглия направляются к а-фазическим нейронам и от них к белым мышцам (быстрым нейромоторным единицам). Импульсация, идущая от веретен по афферентным волокнам группы Iа в спинном мозге моносинаптически возбуждает мотонейроны своей мышцыи через тормозящий интернейронтормозит мотонейроны мышцы-антагониста (реципрокное торможение). Аннуло-спиральные терминали могут также возбуждаться (в ответ на их деформацию) при сокращении интрафузальных мышечных волокон под влиянием нервных импульсов от γ-нейронов (центральное воздействие).
К ядерно-цепочечным волокнам также подходят чувствительные окончания афферентных волокон группы II- (образовано 1-2 тонкими чувствительными нервными волокнами диаметром около 8 мкм.) — так называемые вторичные окончаниякоторые заканчиваются на них гроздьюбляшек — это колечки, розетки, утолщения терминалей.
Они механически деформируются и возбуждаются под влиянием статической длины мышцы. Афферентные волокна группы IIвозбуждают мотонейроны мышц-сгибателей и тормозят мотонейроны мышц-разгибателей. Имеются, однако, данные, что афферентные волокна группы II, идущие от мышц-разгибателей, могут возбуждать мотонейроны своей мышцы.Потенциалы действия, пришедшие к атоническим нейронам по волокнам второй группы, далее направляются к красным мышцам (медленным нейромоторным единицам).
Гамма-регуляция мышечного тонуса. Степень сокращения (напряжения) мышцы зависит от частоты импульсов, приходящих к ней от а-мотонейронов. Частота разрядов а-мотонейронов в свою очередь регулируется импульсами от проприорецепторов этой же мышцы. Веретена имеют и эфферентную иннервацию: интрафузальные мышечные волокна иннервируются аксонами, идущими к ним от γ-мотонейронов. Эти так называемые γ -эфферентные волокна подразделяют на динамические и статические. Разряды γ -мотонейронов повышают чувствительность мышечных веретён, причем динамические γ-эфференты преимущественно усиливают реакцию на скорость удлинения мышцы, а статические — на длину. Активация у-эфферентови без растяжения мышцы сама по себе вызывает импульсацию афферентов веретен вследствие сокращения интрафузальных мышечных волокон. В результате получается, что при одинаковой длине мышцы увеличивается поток импульсов от рецепторов к а-мотонейронам, а от них — к мышце. Тем самым повышается мышечный тонус. Уровень возбуждения γ -системы тем выше, чем интенсивнее возбуждены а-мотонейроны данной мышцы, т. е. чем больше сила ее сокращения. Сами γ -мотонейроны контролируются ретикулярной формацией ствола мозга, мозжечком и корой.
Благодаря сложному строению мышечные веретёна реагируют не только на периферические возбуждения (изменение длины мышцы), но и на изменения активных центральных механизмов — возбуждение γ-мотонейронов («γ-петля»).
Таким образом, веретена можно рассматривать как непосредственный источник информации о длине мышцы и ее изменениях, если только мышца не возбуждена. При активном состоянии мышцы необходимо учитывать влияние у-системы. Во время активных движений γ-мотонейроны поддерживают импульсацию веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на неравномерности движения как увеличением, так и уменьшением частоты импульсации и участвовать таким образом в коррекции движений.
Сухожильные рецепторы Гольджи (тельца Гольджи) находятся в сухожилиях. Это гроздевидные чувствительные окончания, достигающие у человека 2-3 мм в длину и 1 — 1,5 мм в ширину. Тельца Гольджи, располагаясь в сухожилиях, включены относительно скелетной мышцы последовательно, поэтому они возбуждаются при ее сокращении вследствие натяжения сухожилия мышцы. Они имеют высокий порог возбуждения.Рецепторы Гольджи контролируют силу мышечного сокращения — напряжения. Идущие от этих рецепторов афферентные волокна относятся к группе Ib. На спиналь-ном уровне они через интернейроны вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста.Информация от мышечных рецепторов по восходящим путям спинного мозга поступает в высшие отделы ЦНС, включая кору большого мозга, и участвует в кинестезии.
Таким образом, состояние мышцы контролируется двумя типами рецепторов — мышечными веретенами, передающими информацию о длине мышцы и скорости изменения этой длины, и сухожильным рецептором, который сигнализирует о силе мышечного сокращения.
В отличие от мышечных веретён рецепторы Гольджи не имеют собственной эфферентной иннервации. Их импульсы направляются в спинной мозг к вставочным тормозным нейронам. Далее происходит переключение импульсов на а-мотонейроны, которые тормозятся, в результате чего мышца расслабляется. Это называется «аутогенным торможением». Все сухожильные рефлексы начинаются с рецепторов Гольджи.
Коленный рефлекс — это искусственный (сухожильный) рефлекс. Он наступает при растяжении мышечного веретена в ответ на удар молоточком. Коленный рефлекс в отличие от других рефлексов не может наступать с рецепторов Гольджи, так как для их раздражения нужна большая сила, а у мышечного веретена порог возбуждения низкий. От рецепторов возбуждение поступает в Ib, а оттуда на четырёхглавую мышцу бедра, и нога в коленном суставе выпрямляется.
Тельца Фатера-Починипредставляют собой инкапсулированные нервные окончания, локализуются в глубоких слоях кожи, в сухожилиях и связках, реагируют на изменения давления, которое возникает при сокращении мышц и натяжении сухожилий и связок.
Суставные рецепторы. Они изучены меньше, чем мышечные. Известно, что суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя таким образом в системе обратных связей от двигательного аппарата и в управлении им.

Рис.1. Рефлексы растяжения моносинаптический (I, от мышечных веретён, приводит к сокращению той же мышцы) и полисинаптический (II)
· Моносинаптическая дуга. Ia-проприоцептивные нервные волокна, отходящие от мышечного веретена, входят в задний корешок спинного мозга и сразу направляются в передний рог, где и образуют синапсы с a‑мотонейронами, посылающими сигналы к мышце.
· Полисинаптическая дуга дополнительно включает вставочный нейрон. На рис.1. — (II) представлена дуга тормозного рефлекса, возникающего при растяжении сухожильных рецепторов Гольджи.
Проводниковый отдел. Чувствительность кожи и ощущение движения обусловлены проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спинно-таламическому, значительно различающимся по своим морфологическим и функциональным свойствам. Существует и третий путь — латеральный тракт Морина, близкий по ряду характеристик к лемнисковой системе.
Лемнисковый путьна всех уровнях состоит из относительно толстых и быстропроводящих миелинизированных нервных волокон.Первые нейроны этого пути находятся в спинномозговом узле, их аксоны в составе задних столбов восходят к тонкому (ядро Голля) и клиновидному (ядро Бурдаха) ядрам продолговатого мозга, где сигналы передаются на вторые нейроны лемнискового пути. Часть волокон, в основном несущих сигналы от суставных рецепторов, оканчивается на мотонейронах сегментарного спинального уровня. Проприоцептивная чувствительность передается в спинном мозге также по дорсальному спинно-мозжечковому, спинно-цервикальному и некоторым другим путям.
В продолговатом мозге вклиновидном ядре — вторые нейроны проприоцептивной чувствительности. Аксоны этих нейронов образуют медиальную петлюи после перекреста на уровнеолив направляются в специфические ядра таламуса — вентробазальный ядерный комплекс. В этих ядрах концентрируются третьи нейроны лемнискового пути. Их аксоны направляются в соматосенсорную зону коры большого мозга.
Спинно-таламический путь. Этот путь значительно отличается от лемнискового. Его первые нейронытакже расположены в спинномозговом узле,откуда они посылают в спинной мозг медленнопроводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спинно-таламического пути направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спинно-таламического пути лишь частично дают проекции в соматосенсорную зону коры.
Спинно-таламический путь служит в большей мере для передачи температурной, болевой и тактильной чувствительности.
.
7
Соседние файлы в папке Разное
- #
- #
- #
- #
- #
- #
- #
Источник