
Рост растяжением у растений
Содержание статьи
Экология СПРАВОЧНИК
Общий характер растяжения гравитропически индуцированных корней, по-видимому, варьирует в зависимости от вида и экспериментальных условий. Кроме того, в отличие от гравитропизма побегов, где определенный тип дифференциального роста устанавливается после восприятия гравитационного раздражения и этот же тип роста сохраняется в период ответной реакции, при гравитропизме корней типы дифференциального роста со временем изменяются. Например, вначале может наблюдаться замедление роста нижней стороны и ускорение роста верхней, но позднее скорость удлинения верхней стороны может снизиться до уровня даже более низкого, чем до помещения корня в горизонтальное положение. Было обнаружено, что у ряда видов изменение ориентации из вертикальной в горизонтальную вызывает снижение общей скорости роста растяжением. Вплоть до недавнего времени общие представления о механизме передачи раздражения при гравитропизме корней основывались на механизме, предложенном Холодным и Вентом в 20-х годах для фототропизма и гравитропизма колеоптилей. Иными словами, -считалось, что ауксин, синтезирующийся или высвобождающийся в апексе корня, подлежал латеральному перемещению книжной стороне органа. Согласно этой теории, верхняя сторона горизонтально помещенного ортогравитропического корня должна содержать близкий к оптимальному уровень ауксина и поэтому расти быстрее, чем ингибируемая нижняя сторона, вследствие чего образуется положительный изгиб корня. Тем не менее вопрос остается открытым, поскольку существуют экспериментальные данные, показывающие, что латеральное перемещение ауксина вниз может происходить у гравитропически индуцированных корней.[ …]
Как мы видели в первой главе этой книги, рост растений в основном является следствием вакуолизации клеток, переходящих из меристемы в зону растяжения. Наиболее ярко это положение иллюстрирует рост растяжением стеблей, колеоптплей, черешков и корней. Поэтому регуляция роста растений фитогормонами в значительной степени определяется их влиянием на процессы клеточного растяжения. Вследствие этого, а также из-за удобства работы с такими удлиняющимися структурами, как колеоптили или стебли, большая часть исследований по механизму действия фитогормонов посвящена явлению роста растяжением.[ …]
Конечно, при изучении действия гормонов на рост растяжением использовали разные подходы. Одиако общей чертой, пожалуй, всех без исключения работ такого типа было удаление эндогенного источника изучаемого гормона (например, путем вырезания сегментов стеблей или колеоптилей) и последующее введение экзогенного гормона. Удаление природного гормона приводит к изменению скорости роста растяжением (к снижению или повышению ее в зависимости от типа гормона), а добавление экзогенного гормона частично или полностью восстанавливает исходную скорость роста. Следовательно, изучаемый эффект гормона является чисто количественным. Это означает, что не следует думать о гормональной индукции каких-то новых типов метаболической активности, например об изменении характера белкового синтеза, хотя, конечно, гормон может и обычно влияет на скорости синтеза белков, что приводит к различиям в скорости роста разных клеток.[ …]
Несомненно, что для нормального длительного роста растяжением необходим непрерывный синтез белка, даже если краткосрочные (с лаг-периодом порядка 10 мин или меньше) ростовые ответы на гормон наблюдаются и без синтеза новых РНК и белков. В случае регуляции роста растяжением ауксином очевидно, что при быстром ростовом ответе происходит выделение И+ в оболочку, а снижение pH или непосредственно ослабляет какие-то межмолекулярные связи, или благоприятствует активности определенных ферментов, разрыхляющих оболочку.[ …]
| Функционирование аппарата Гольджи, приводящее к росту растяжением кончика гифы гриба | 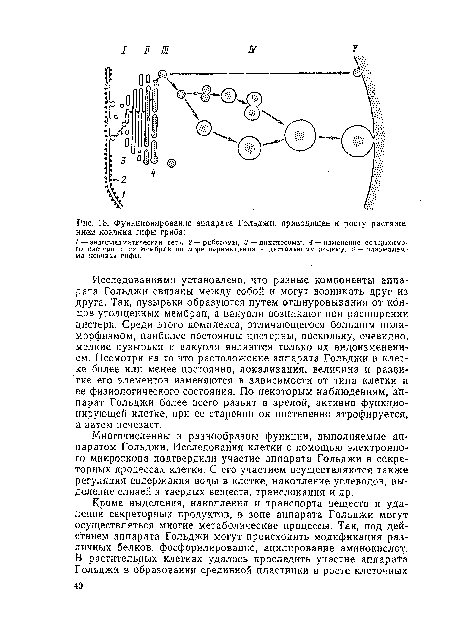 |
Утолщение стеблей и корней происходит за счет деления клеток, роста растяжением и дифферепцировки. У двудольных растений и у большинства голосеменных радиальный рост стеблей и корней обеспечивается активностью латеральной меристемы. Обычно различают два типа латеральной меристемы: сосудистый камбий и пробковый камбий, или среллоген. У тех видов-однодольных растений, которые в какой-то степени растут в толщину, клетки камбия также играют определенную, хотя и более ограниченную роль.[ …]
После заложения листа в апексе побега начинаются процессы его роста и развития, включающие клеточное деление, рост, растяжением и диффёренцировку (см. гл. 2). Естественно думать, что эти процессы находятся под контролем фитогормонов, одним из которых, очевидно, является ауксин. Однако нельзя сказать, что действие ауксина связано со всеми аспектами роста листа. Было обнаружено, что ауксины в зависимости от их концентрации могут стимулировать или ингибировать рост центральной и боковых жилок, но мало влияют на ткани мезофилла между жилками. В настоящее время гормональная регуляция роста листа изучена мало. Известно только, что ауксин, по-видимому, необходим для роста жилок.[ …]
Ауксины и гиббереллины взаимодействуют не только при регуляции роста растяжением, но и во многих других случаях. Например, комбинация ауксина и гиббереллина часто более эффективно, чем каждый из них в отдельности, вызывает пар-теиокарпию плодов или усиливает активность камбия (с. 187).[ …]
Как известно, круговые нутации обусловлены различными скоростями роста растяжением на разных сторонах органа. Для проявления круговой нутации скорости удлинения сторон по окружности органа должны ритмически изменяться, отчего и возникает раскачивание растения. Для объяснения круговой нутации было предложено несколько механизмов, в том числе: а) автономные колебательные изменения концентраций эндогенных ростовых гормонов вокруг оси органа, б) «перекомпенса-ция» геотропической реакции, в) комбинации автономных процессов и геотропической перекомпенсации.[ …]
Начальное снижение прироста корней связано с нарушением деления меристематических клеток, но рост растяжением сохраняется. Последний прекращается только при больших дозах воздействия, вызывающих гибель клеток.[ …]
Кроме того, этилен регулирует рост в длину у водных растений, например у риса, Са1Шг1сНе и Ыутркаеа. Эта стимуляция продолжается до тех пор, пока растущие части растения не достигнут поверхности воды и не смогут выделить этилен в атмосферу: погруженные в воду побеги (и листовые черешки) очень медленно отдают свой этилен в окружающую среду из-за плохой растворимости этого газа в воде. Таким путем растения приспосабливаются к высоте уровня воды.[ …]
Ауксины и гиббереллины могут при некоторых условиях стимулировать деление клеток, однако вх скорее можно назвать гормонами, регулирующими рост растяжением. Было показало, что клетки каллюса в стерильной культуре через определенный промежуток времени прекращают деление. Однако при добавлении к питательной среде производных ДНК, получающихся после ее авто-клавнрования, деление клеток возобвовляется.[ …]
Относительно высокие интенсивности излучения рассеянного синего света оказывают быстрое ингибирующее действие (латентный период менее 5 мин) на рост растяжением у зеленых стеблей. Величина ингибирующего эффекта синего света зависит от уровня излучения, но на продолжительность латентного периода уровень излучения не оказывает влияния. Эти особенности действия синего света на рост растяжением у неэтиоли-рованных растений логически привели к недавно высказанному предположению, что фототропизм, по-видимому, является результатом прямого ингибирующего действия синего света на рост клеток растяжением на той стороне органа, которая подвергалась освещению. Пигментная система (возможно, флаво-протеид), обусловливающая эффекты .синего света и явление фотоморфогенеза, индуцируемого световым излучением высокой энергии, вероятно, одна и та же (см. гл. 8). Фоторецептор синего света иногда называют криптохромом. Быстрота действия синего света наводит на мысль, что различия в транспорте гормонов не играют никакой, роли в явлении фототропизма. Если окажется, что прямая реакция на синий свет лежит в основе фототропизма растений, то сходство этого механизма со значительно лучше установленным механизмом фототропизма у спо-рангиеносцев фикомнцетов (с. 277) очевидно, и этот факт представляется крайне интересным.[ …]
Относительно высокие интенсивности излучения рассеянного синего света оказывают быстрое ингибирующее действие (латентный период менее 5 мин) на рост растяжением у зеленых стеблей. Величина ингибирующего эффекта синего света зависит от уровня излучения, но на продолжительность латентного периода уровень излучения не оказывает влияния. Эти особенности действия синего света на рост растяжением у неэтиоли-рованных растений логически привели к недавно высказанному предположению, что фототропизм, по-видимому, является результатом прямого ингибирующего действия синего света на рост клеток растяжением на той стороне органа, которая подвергалась освещению. Пигментная система (возможно, флаво-протеид), обусловливающая эффекты .синего света и явление фотоморфогенеза, индуцируемого световым излучением высокой энергии, вероятно, одна и та же (см. гл. 8). Фоторецептор синего света иногда называют криптохромом. Быстрота действия синего света наводит на мысль, что различия в транспорте гормонов не играют никакой, роли в явлении фототропизма. Если окажется, что прямая реакция на синий свет лежит в основе фототропизма растений, то сходство этого механизма со значительно лучше установленным механизмом фототропизма у спо-рангиеносцев фикомнцетов (с. 277) очевидно, и этот факт представляется крайне интересным.[ …]
Углы, под которыми листья и боковые ветви отходят от стебля у двудольных и голосеменных, определяются относительными (дифференциальными) скоростями роста растяжением внутренней (адаксиальной) и наружной (абаксиалы-юй) сторон органов. В качестве примера эпинастии листьев можно привести ростовой изгиб черешка: верхняя (адакснальная) сторона удлиняется быстрее, чем нижняя (абаксиальная), в результате чего по мере роста листовая пластинка движется вниз (рис. 7.1). При гипонастии (движении кверху) более быстрым ростом характеризуется абаксиальная сторона органа. Эпинастические движения листа происходят за счет ростового изгиба, который по мере развития листа распространяется базипетально от основания листовой пластинки вдоль черешка.[ …]
В основных изученных типах органов побега (колеоптилях, гипокотилях и междоузлиях) отрицательный гравитропический изгиб обычно возникает в результате замедления или полного прекращения роста растяжением всей верхней стороны и ускорения роста — нижней. У зрелых злаков междоузлия при вертикальном положении стебля не удлиняются вообще, но при горизонтальном расположении побега нижняя сторона начинает удлиняться, изгибаясь кверху, а клетки верхней стороны сдавливаются. Общие закономерности роста при гравитропизме корней более разнообразны и сложны и будут рассмотрены позднее (с. 293).[ …]
Одним из важнейших факторов, влияющих и регулирующих индукцию покоя у древесных растений, является длина дня. У большинства видов, изученных в этом отношении, длинные дни ускоряют вегетативный рост, а короткие дни вызывают прекращение роста растяжением и образование зимующих почек у сеянцев древесных растений (рис. 11.2). Однако ряд обычных культивируемых плодовых деревьев (груша, яблоня, слива) и некоторые другие виды, в том числе представители семейства маслинных, по-видимому, относительно нечувствительны к изменениям длины дня.[ …]
Хорошо известно, что для широко распространенных видов древесных растений, таких, как Pinus silvestris, Picea abies, в отношении фотопериодической реакции характерны заметные экотипические различия в зависимости от широты и высоты места произрастания. Северным расам для активного роста растяжением требуются более длинные фотопериоды, чем более южным расам, адаптированным к более коротким естественным фотопериодам. Этот факт наводит на мысль, что древесные растения лучше приспособлены к естественным условиям длины дня и что последние, видимо, играют важную регулирующую роль в сезонном цикле роста и покоя.[ …]
Было установлено, что сезонное уменьшение длины дня играет важную роль в детерминации наступления покоя у сеянцев тех видов, которые обычно продолжают активный рост осенью, например Larix decidua, Poputus spp., Robinia pseudacacia. Правда, очень часто можно обнаружить, что старые растения имеют намного более короткий период роста растяжением, чем сеянцы тех же видов, и прекращают рост уже в нюне или июле, когда естественные фотопериоды еще достаточно длинные. В таком случае сомнительно, чтобы сокращение длины дня играло важную роль в детерминации образования покоящихся почек, и более вероятно, что какие-то изменения в содержании питательных веществ или в гормональном балансе, возникающие внутри самого растения, определяют период роста и наступления покоя. Однако, как мы видели, зимующие почки вначале находятся в состоянии нредпокоя и только потом входят в глубокий покой; возможно, что сокращение длины дня осенью играет роль в переходе гточек от предпокоя к глубокому покою.[ …]
По устойчивости против выпревання разные Гибридные растения, отрастающие короткими генеративными побегами, вели себя неодинаково. Наряду с растениями, плохо переносящими мягкие многоснежные зимы, встречаются растения, устойчивые-против выпревания. Весной эти растения рано и быстро трогаются в рост растяжением стебля, и у них очень рано начинает формироваться колос.[ …]
Характерной: чертой ростовых процессов растительных организмов является их локализация в определенных тканях — меристемах. Клетки меристемы делятся, дочерние клетки достигают размеров материнской и снова делятся. Это связано с тем, что большинство меристемати-ческих клеток через несколько делений (4—5) переходят к росту растяжением. Однако есть клетки, которые делятся в течение всей жизни органа. Такам образом, меристемы порня и стебля состоят из двух тисов клеток, резко отличающихся по функциям и по способности к делению. Меристематические клетки, расположенные па самом верху стебля или корпя, не прекращают деления в течение всего периода роста. Более длительная способность к делению является следствием меиыпей частоты делений в большей длительности питерфазы. Одновременно данные клеткл характеризуются в большей длительностью митотического цикла. Вместе с тем эти клетки более устойчивы к неблагоприятным воздействиям. Так, в них реже возникают хромосомные аберрации, что очень важно для сохранения жизнеспособности организма (В. Б. Иванов) Клетки меристемы ожидания стебля менее дифференцированы, что облегчает их последующую дифференциацию и имеет значение при переходе к образованию генеративных органов.[ …]
У большинства травянистых растений, используемых п вирусологических экспериментах, отдельные листья никогда не находятся в стационарном состоянии. Они обычно проходят через четыре стадии, плавно переходящие одна в другую. Первая стадия, в ходе которой лист табака, например, достигает около 2 см в длину, — это стадия интенсивного клеточного деления. Ее сменяет стадия, иа которой рост (растяжение) клеток и синтез белка становятся преобладающими. По мере того как лист приближается к своим окончательным размерам, самым активным процессом становится фотосинтез (он продолжается с самой ранней стадии), и, наконец, на четвертой стадии берут верх процессы старения.[ …]
В ранних работах высказывались предположения о прямом влиянии ауксина на клеточную стенку, в частности на ионные мостики между входящими в ее состав полиуронидами («пектиновыми веществами») (рис. 4.12), но позднее такая точка зрения была пересмотрена. Тем не менее не так давно было обнаружено, что полиурониды, возможно связанные с клеточной стенкой, могут служить рецепторами для гиббереллинов при стимуляции последними роста растяжением гипокотилей салата-латука (см. с. 130).[ …]
Подводя итог, следует сказать, что, по имеющимся данным, при освещении верхушек этиолированных колеоптилей низкими дозами одностороннего света в первую очередь происходит восприятие светового раздражения, а затем следует поперечное перемещение молекул эндогенного ауксина в пределах верхушки. В случае положительной фототропической реакции ауксин движется к затененной стороне, т. е. больше ауксина перемещается в зону реакции, расположенную под более темной половиной верхушки, и как следствие этого происходит усиленный рост растяжением в этой области, в результате чего весь колсо-птиль изгибается по направлению к источнику света.[ …]
Скорость акролетального передвижения ауксина в надзем-. ных органах не была точно измерена, но обычно она гораздо меньше, чем скорость базипетального передвижения. Однако ряд факторов влияет на соотношение скоростей базипетального и акропетального транспорта. Так, полярность движения ауксина снижается с увеличением возраста изучаемой ткани. Пока неясно, происходит ли это за счет снижения базипетального передвижения, за счет усиления акропетального тока или за счет обоих этих явлений. Тем не менее очевидно, что процессы созревания в тканях связаны с постепенным снижением полярности транспорта ауксина. В связи с этим было высказано предположение, что полярный транспорт ауксина происходит только в процессе роста клеток растяжением. Вместе с тем ранее было обнаружено, что ГАз стимулировала базипетальное передвижение ИУК в тканях стебля. Таким образом, мы не знаем ни то, каким образом происходит уменьшение полярности передвижения ауксина в зрелых тканях, ни то, какое это может иметь значение.[ …]
Источник
рост растяжением
рост растяжением
ЭМБРИОЛОГИЯ РАСТЕНИЙ
РОСТ РАСТЯЖЕНИЕМ – фаза в развитии растительной клетки, в ходе которой размеры клетки увеличиваются за счет образования и последующего увеличения центральной вакуоли. Накапливающиеся в вакуоли ионы и нейтральные молекулы способствуют осмотическому поступлению в вакуоль воды. Одновременно увеличивается эластичность и растяжимость клеточной стенки, снижается противодавление клеточной стенки на клетку.
Общая эмбриология: Терминологический словарь — Ставрополь.
О.В. Дилекова, Т.И. Лапина.
2010.
Смотреть что такое «рост растяжением» в других словарях:
РОСТ РАСТЯЖЕНИЕМ — увеличение размеров вновь образовавшихся клеток, наблюдаемое обычно после неоднократного предварительного их деления … Словарь ботанических терминов
ДВИЖЕНИЕ — у растений, перемещения растения или его частей в пространстве. Растения, как правило, ведут прикреплённый образ жизни, поэтому их двигат. активность выражается в подвижности цитоплазмы и органоидов клеток, а также в перемещении их органов.… … Биологический энциклопедический словарь
ВОДНЫЙ РЕЖИМ РАСТЕНИЙ — сово купность процессов поглощения, усвоения и выделения воды растениями. Вода, составляющая 80 95% массы растения, является средой для биохимич. реакций, участвует в фотосинтезе, обеспечивает структуру коллоидов цитоплазмы, опре дел. конформацию … Биологический энциклопедический словарь
Ауксины — * аўксіны * auxins растительные гормоны, которые индуцируют рост клеток и их деление, способствуют аттрагирующему действию тканей, активируют протонную помпу, регулируют рост растяжением и устьичные движения. Естественным А. является производное… … Генетика. Энциклопедический словарь
СЕРДЦЕ — СЕРДЦЕ. Содержание: I. Сравнительная анатомия……….. 162 II. Анатомия и гистология……….. 167 III. Сравнительная физиология………. 183 IV. Физиология………………. 188 V. Патофизиология……………. 207 VІ. Физиология, пат.… … Большая медицинская энциклопедия
ГОРМОНЫ РАСТЕНИЙ — или фитогормоны, вырабатываемые растениями органические вещества, отличные от питательных веществ и образующиеся обычно не там, где проявляется их действие, а в других частях растения. Эти вещества в малых концентрациях регулируют рост растений и … Энциклопедия Кольера
ПРЕДЕЛ ТЕКУЧЕСТИ — в сопротивлении материалов, напряжение, при к ром начинает развиваться пластич. деформация. В опытах с растяжением цилиндрич. образца определяется нормальное напряжение ss в поперечном сечении, при к ром впервые возникают пластич. (необратимые)… … Физическая энциклопедия
Грудной ребёнок — I Грудной ребёнок ребенок в возрасте до одного года. Выделяют период новорожденности, продолжающийся 4 нед. после рождения (см. Новорожденный (Новорождённый)) и грудной возраст (от 4 нед. до 1 года). В грудном возрасте ребенок растет и… … Медицинская энциклопедия
Грудной ребёнок — Рис. 1. Схематическое изображение проекций большого и малого родничков. Рис. 1. Схематическое изображение проекций большого (а) и малого (б) родничков. Грудной ребёнок ребенок в возрасте до одного года. В первые 4 нед. жизни его называют… … Первая медицинская помощь — популярная энциклопедия
ауксины — группа гормонов растений. Регулируют на разных этапах жизни растения его рост, дифференцировку органов, ростовые реакции на свет и силу тяжести. По химической природе производные индола. Основной представитель индолилуксусная кислота. * * *… … Энциклопедический словарь
Книги
- Биогенез растительных волокон, Агеева М. В., Горшкова Т. А., Аменицкий С. И.. В монографии обобщены представления о биогенезе растительных волокон как индивидуальных клеток, характеризующихся исключительной длиной и мощно развитой клеточной стенкой. С фундаментальной… Подробнее Купить за 789 грн (только Украина)
- Биогенез растительных волокон, Горшкова Т.А.. В монографии обобщены представления о биогенезе растительных волокон как индивидуальных клеток, характеризующихся исключительной длиной и мощно развитой клеточной стенкой. С фундаментальной… Подробнее Купить за 738 руб
- Биогенез растительных волокон, Агеева М. В., Горшкова Т. А., Аменицкий С. И.. В монографии обобщены представления о биогенезе растительных волокон как индивидуальных клеток, характеризующихся исключительной длиной и мощно развитой клеточной стенкой. С фундаментальной… Подробнее Купить за 616 руб
Источник